Arq. Bras. Oftalmol. 2019; 82 (6): 10.5935/0004-2749.20190103
Total: 2174
Ana Cláudia Viana Wanzeler1; Italo Antunes França Barbosa1; Bruna Duarte1,2; Daniel Borges1; Eduardo Buzolin Barbosa1; Danielle Kamiji1; Delma Regina Gomes Huarachi1; Mônica Barbosa de Melo1,3; Mônica Alves1
DOI: 10.5935/0004-2749.20190103
ABSTRACT
Pterygium pathogenesis has been mainly asso ciated with UV light exposure; however, this association remains quite controversial. The complete mechanism of pterygium also remains to be clarified. Factors such as inflammation, viral infection, oxidative stress, DNA methylation, inflammatory mediators, extracellular matrix modulators, apoptotic and oncogenic proteins, loss of heterozygosity, microsatellite instability, lymphangiogenesis, epithelial-mesenchymal cell transition, and alterations in cholesterol metabolism have been identified as causes. Several studies aimed to clarify the molecular mechanisms underlying the growth and proliferation of pterygium. Understanding its molecular basis provides new potential therapeutic targets for its prevention and treatment. A comprehensive search of the databases, namely, MedLine, EMBASE, and LILACS, was conducted with the following key words: pterygium, epidemiology, pathogenesis, biomarkers, and review. This review describes the epidemiology, clinical presentation, and current investigation of biological mediators involved in pterygium development.
Keywords: Pterygium/epidemiology; pathogenesis; Biomarkers; Review
RESUMO
A patogênese do pterígio tem sido relacionada, prin cipalmente, à exposição à luz ultravioleta, mas esta asso ciação permanece bastante controversa. O mecanismo completo do pte rígio também permanece por esclarecer. Fatores como inflamação, infecção viral, estresse oxidativo, metilação do DNA, mediadores inflamatórios, moduladores de matriz extracelular, proteínas apoptóticas e oncogênicas, perda de heterozigose, instabilidade de microssatélites, linfangiogênese, transição celular epitelial-mesenquimal e alterações no metabolismo do colesterol tem sido identificados como causas. Diversos estudos visam esclarecer os mecanismos moleculares subjacentes ao crescimento e proliferação do pterígio. Entender sua base mo lecular fornece novos alvos terapêuticos potenciais para sua prevenção e tratamento. Uma busca abrangente nas bases de dados, a saber, MedLine, EMBASE e LILACS, foi realizada com as seguintes palavras-chave: pterígio; epidemiologia; patogênese; biomarcadores e revisão. Esta revisão descreve a epidemiologia, apresentação clínica e a atual investigação de mediadores biológicos envolvidos no desenvolvimento do pterígio.
Descritores: Pterígio/epidemiologia; patogênese; Biomarcadores; Revisão
INTRODUCTION
Pterygium is a nonneoplastic elastotic degeneration of subepithelial growth, originating from the bulbar conjunctiva that extends to the corneal surface, and even reached the visual axis in some cases. It is a common ocular surface disorder, especially in geographical areas near the equator. The exact cause of pterygium remains unclear; however, some risk factors are identified as causes, with long-term ultraviolet radiation exposure as the most important1,2. Although pterygium is generally regarded as a benign and cosmetic problem, it may result in significant visual morbidity or even potential blindness in severe cases if not properly treated3. It usually occurs in the nasal area, but can develop temporally or in both directions and may even occur bilaterally. Pterygium surgery is generally considered when symptoms do not respond to conservative treatment, when it induces visual disturbances, disability, or for cosmetic purposes4,5.
With regard to the mechanisms, genetic factors were suggested in some studies, noting that some genes associated with DNA repair play a crucial role in pterygium development. However, studies on genetic variant contributions are limited in sample size and should be cautiously interpreted. Chronic irritation with actinic damage is likely responsible for the typical fibrovascular reaction of pterygium6. Growth factors, cytokines, and matrix metalloproteinase are involved in the pathogenesis of pterygium and, along with UV exposure, may trigger the proinflammatory aspects7.
This review describes the epidemiology, clinical presentation, and investigation of biological modulators found in recent literatures. Therefore, the following keywords were searched: pterygium, epidemiology, angiogenesis, proliferation, inflammation, gene, protein, pathogenesis, and tight junction proteins.
Epidemiology
The prevalence of pterygium has been investigated in several population-based studies. Rates widely vary depending on the studied population, ranging from 2.8% to 38.7%, as recent studies in South Korea and China found prevalences of 8.8%8 and 9.84%9, in contrast to 16% in Arizona (USA)10 and 38.7% in Northwest Ethiopia11.
In Brazil, no population data were conducted on pterygium prevalence throughout the country, and only a few studies investigated this subject even for specific regions. A survey in riverside communities at the Solimoes and Japura rivers, in Amazonas, showed that its prevalence in the general population was 21.2%12. Another survey, still in the Brazilian Amazon, reported that up to 36.6% prevalence of pterygium was observed in the indigenous population13. In the southeast region, a study in Botucatu City revealed a prevalence of 8.12%14.
A large-scale survey on the rate of pterygium and other ocular diseases has not been conducted yet. A meta-analysis published in 2013, covering a total of 20 studies involving 12 countries with 900,545 samples, showed a combined pterygium prevalence rate of 10.2% (95% confidence interval [CI]: 6.3% to 16.1%), in the general population15. Table 1 summarizes the epidemiological information.
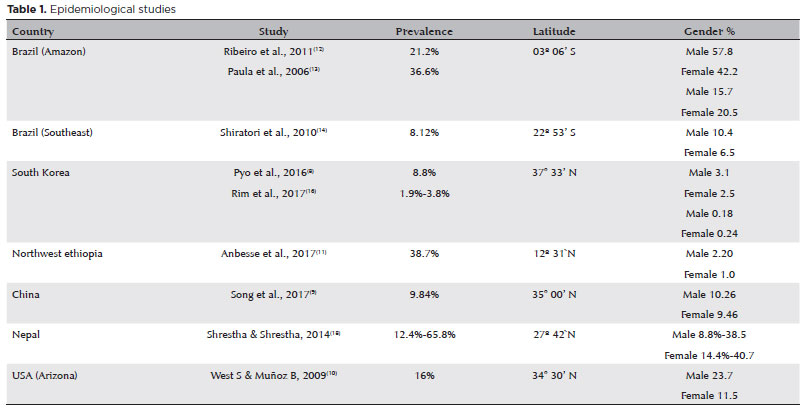
Pterygium more frequently occurs in young adults, rarely before aged 15 years. It is believed to have some inherited patterns; however, several risk factors have been reported, such as dust exposure, heat, inflammation, and eye surface infection, rural residency, advanced age, low educational levels, and outdoor activity16.
The prevalence of pterygium has been described as higher in males16; however, other studies have shown that both genders have the same proportions17 or even predominantly occurred in women18. Its relationship with smoking was also investigated, but remains inconclusive15.
Clinical presentation
Pterygium is a conjunctival fibrovascular tissue that extends to the cornea and can lead to irritating symptoms, visual disturbance, recurrent inflammation, and aesthetic alterations in the ocular surface. Its diagnosis is confirmed with slit-lamp examination, in which the pterygium can be classified as grade 1, when the fibrovascular tissue reaches the limbus; grade 2, when it covers the cornea in approximately 2 mm; grade 3, when it reaches the pupil margin; and grade 4, when it exceeds the pupil (Figure 1). As regards its morphological features, pterygium can be classified as involutive or atrophic when it allows visualization of structures immediately below the lesion and as inflamed once the fibrovascular tissue is fleshy and prevents visualization of the structures below19,20. Recently, a functional classification using the corneal topographic data based on corneal higher-order irregularity was proposed by Miyata et al. to objectively evaluate pterygium severity. Hence, pterygium was graded based on corneal irregularity within the three zones: 1.0, 3.0, and 5.0 mm diameters. Thus, increased corneal irregularity within a 1.0-mm diameter was considered to highly result in the risk for visual function impact, and increased corneal irregularity within a 5.0-mm diameter was considered as mild severity with visual function influences21.

Visual impairment induced by pterygium growth occurs due to induced astigmatism and opacification in the visual axis, requiring surgical treatment, as well as recurrent inflammation that does not improve with topical treatment20.
Its surgical procedure consists of dissecting the head of the pterygium from the cornea and resection of the conjunctiva and Tenon's capsule. Several surgical techni ques have been used, with excision of the pterygium followed by autologous conjunctival grafts as the most common, showing lower recurrence rates. Recurrence after a surgical treatment can be identified by the growth of conjunctival vessels toward the limbal edge inducing fibrous tissue growth into the cornea and representing poor outcomes22. However, clinical features such as extensive size, inflammation, and recurrent lesions remain challenges during the surgical treatment23,24.
Pathogenesis
UV radiation exposure
UV radiation from the sunlight is divided into three categories:
1. UVA (wavelength, 320-400 nm) has the longest wavelength and maximum penetration power; thus it is not attenuated by the ozone layer. Is an important inducer of pigmentation and contributes to premature skin aging, immunosuppression, and carcinogenesis25.
2. UVB (wavelength, 280-320 nm) is absorbed by the ozone layer and comprises approximately 1%-10% of the total UV radiation that reaches the Earth's surface. It is responsible for various biological events, including sunburn, immunosuppression, and carcinogenesis25.
3. UVC (wavelength, 200-280 nm) has the highest energy among the three UV rays and possesses strong mutagenic properties. It is almost completely absorbed by the ozone layer, thereby imposing negligible effects to human eyes26.
UVB light exposure has been attributed as a major cause of pterygium. This kind of radiation can potentially harm and alter cells and tissues through direct phototoxic effects on the cellular DNA and generation of reactive oxygen species, which damage the cellular DNA. Wavelengths below 300 nm have been known as the most biologically active forms and are absorbed by the cornea. Exposure to UVB radiation causes oxidative stress, which may lead to upregulation of many potential mediators of pterygium growth27-29 as shown in figure 2.

Viral infections
The polymerase chain reaction technique allowed examination of the alleged involvement of viral infections in the process of pterygium pathogenesis. Some reports demonstrated the presence of herpes simplex virus and human papilloma virus (HPV) in pterygium samples30,31. Viruses encode proteins that inactivate p53, leading to chromosomal instability and increasing the likelihood of cell progression to malignancy. HPV is most frequently found in the pterygium, with variable pre valence rates32. Its involvement as a cofactor in the pterygium pathogenesis is suggested, but remains controversial. If indeed HPV is involved in pterygium pathogenesis or recurrence, anti-viral medications or vaccination may be new options in pterygium therapy30,33.
Molecular mechanisms
Many studies have proposed possible mechanisms of pterygium development, including oxidative stress, extracellular matrix modulators, apoptotic and oncogenic proteins, loss of heterozygosis, DNA methylation, inflammatory mediators, lymphangiogenesis, transition from mesenchymal epithelial cells, and cholesterol metabolism alterations. These studies show evidences that several molecules, such as matrix metalloproteinases (MMPs), growth factors, and interleukins (ILs), are related to proliferation, inflammation, angiogenesis, and fibrosis, as shown in figure 3 and detailed below34-36.

Tumor suppressor genes
Tumor suppressor genes prevent cells from converting into cancer cells and regulate cell growth along with proto-oncogenes37. One of the tumor suppressor genes that have been extensively studied is p53. A survey38 showed that >20% of all pterygium samples were positive for p53 expression. Another immunohistochemical study37 evaluated 13 pterygium samples and 2 normal conjunctiva samples, which showed that 54% of pterygium were positive for p53 aberrant expression, whereas no pathological staining was observed in the normal conjunctiva. Therefore, the aberrant expression of p53 is suggested to promote cell proliferation and slow down apoptosis, thereby accelerating the development of pterygium; besides, the possible growth of limbal tumors is also suggested to be caused by cellular DNA damage that causes mutations in other genes39. In addition to p53, other tumor suppressor genes, such as p63, p16, and p27, were possibly involved in the development of pterygium. P63 is more expressed in the basal and parabasal layers in primary pterygium and in the total thickness of the epithelium in recurrent pterygium. Increased expression of p16 protein was also observed in pterygium. Both p63 and p16 appeared to be rarely expressed in the normal conjunctiva40. P27 gene showed low nuclear immuno reactivity in pterygium tissues, differing from other tumor suppressor genes41.
Apoptosis-related proteins
Survivin is a protein encoded by the BIRC5 gene in humans; it is a member of the apoptosis inhibitory gene family and is expressed in the pterygium epithelium42. The molecular mechanisms of survivin regulation are still not well understood; however, survivin regulation seems to be associated with the p53 protein. Oxidative stress has been demonstrated to be caused by the acti vation of survivin leading to pterygium growth43. In addition, survivin has been found to be highly expressed in all pterygium tissues, but not in the normal human conjunctiva. Survivin was found to be closely related with COX-2 in primary pterygium, suggesting an antiapoptotic mechanism44.
Bcl-2 is the founding member of the Bcl-2 family of apoptosis regulatory proteins, which can induce or inhibit apoptosis. It is encoded by the Bcl-2 gene in humans45,46. Bcl-2 expression was noted in the basal epithelial layer of all pterygium epithelial cells, whereas the normal conjunctiva showed no evidence of the protein39. Decreased miR-122 expression in the pterygium can result in cell apoptosis abnormalities due to its regulation of Bcl-w expression, also a gene of the Bcl-2 family, anti-apoptotic, and subsequently contribute to the development of pterygium47.
Rapamycin complex 1 (mTORC1) is a central regulator of cell growth, proliferation, protein synthesis, autophagy, and transcription. The role of mTORC1 is to activate the protein translation. mTOR signaling is highly activated; therefore, aberrant apoptosis and cell proliferation were observed in pterygium samples. Activation of mTORC1 has been shown to inhibit apoptosis in pterygium by regulating Beclin-1-dependent autophagy by targeting Bcl-2. mTORC1 also negatively regulates the fibroblast growth factor receptor 3 (FGFR3) through the inhibition of p73, thereby stimulating cell proliferation in pterygium. This demonstrates that mTORC1 signaling is highly activated in pterygium and provides new pathways on its pathogenesis and progression48.
Cell adhesion molecules
Cell adhesion molecules play an important role in various physiological and pathological phenomena. These proteins are located on the cell surface and are intrinsically involved in cell binding and other extracellular matrix related to cell adhesion, including selectin, and integrin49. The expression of intercellular adhesion molecule-1 (ICAM-1) is found to be present in pterygium and absent in the epithelium of a normal conjunctiva50. E-cadherin and beta-catenin have also been suggested to be concentrated in the pterygium tissue and are possibly involved in the epithelial proliferation and adhesion51.
Proliferation-related proteins
Proliferation-related proteins such as Ki-67, cyclin D1, and nuclear proliferation antigen play a key role in the cell cycle. Ki-67 is an important marker of cell proliferation. An abnormal expression of ki-67 was found in pterygium samples when compared to a normal conjunctiva52. Proliferating cell nuclear antigen (PCNA) is a nuclear nonhistone protein necessary for DNA synthesis, and its expression may be used as a marker of cell proliferation. The expression of PCNA was significantly higher in pterygium than that in a normal conjunctiva53. Cyclin D1 is a well-known cell cycle control gene that promotes cell cycle progression. A study found that PCNA and cyclin D1 were overexpressed in the limbal part of pterygium epithelial cells as compared with normal conjunctiva samples, which might lead to hyperproliferation of epithelial cells54. Cyclin D1 protein expression in fleshy pterygium was found to be significantly higher than that in the atrophic ones. Another study indicated that b-catenin expressed in the nuclei/cytoplasm could increase cyclin D1 protein expression, which favors the proliferation of pterygium cells55.
Heat shock proteins
Heat shock proteins (HSP) are a protein family produced by cells in response to exposure to stressful conditions. They were first described in relation to heat shock, but are recently known to be expressed during other stresses, including exposure to cold temperatures, UV light, and during wound healing or tissue remodeling52. The expression of HSPs, i.e., Hsp27, Hsp70, and Hsp90, and hypoxia-inducible factor-1α (HIF-1α) were increased in pterygium. The expression of Hsp27 was detected in the epithelial, endothelial, and vascular smooth muscle cells in pterygium, but only in the epithelium in normal conjunctiva56. Changes in HIF-1α and HSP levels in pterygium are believed to represent an adaptive process for cell survival under stressful conditions57.
Tight junction proteins
Tight junction proteins represent a form of cell-to-cell adhesion in the epithelial or endothelial cell layers, forming continuous seals around the cells and also serving as a physical barrier to prevent solutes and water from passing freely through the paracellular space. Claudin family proteins are an important part of this functional and structural barrier and dysregulation on its expression may result in various diseases including cancer58. In normal cornea and conjunctiva, claudin-1 and claudin-4 positivity were demonstrated immunohistochemically59.
Claudins are indispensable proteins for the formation and maintenance of tight junctions. A strong immunohistochemical expression of claudin-1 was found in epithelium conjunctiva samples, whereas its expression in the pterygium samples was low. The significant decrease in claudin-1 expression in the pterygium compared to the normal conjunctiva seems to be involved in the pathogenesis of pterygium60.
Extracellular matrix proteins
The extracellular matrix (ECM) is a collection of ex tracellular molecules secreted by support cells that provide structural and biochemical support to the surrounding cells61.
The aberrant expression of extracellular matrix proteins is believed to may be directly associated with the proliferative growth of pterygium, because it is a fi brovascular tissue characterized by an excessive deposition of extracellular matrix and vascular growth. The extracellular matrix proteins contain keratin, elastin, collagen, and fibrin, among others. K8, K16, K14, and AE3 have been known to be present throughout the thickness in the pterygium epithelium but are absent in the normal conjunctiva62. In fact, pterygium samples showed a higher mRNA level and tropoelastin expression than the conjunctival tissue. Type II collagen expression was positive only in pterygium, whereas collagen types I, III, and IV were detected in both the pterygium and normal conjunctiva63.
Matrix metalloproteinases and tissue inhibitors of metalloproteinases
Matrix metalloproteinases (MMPs), also known as ma trixins, hydrolyze components of the extracellular matrix. These proteinases play a central role in several biological processes, such as embryogenesis, normal tissue remodeling, wound healing, and angiogenesis, and in diseases such as atheroma, arthritis, cancer, and tissue ulceration64. MMPs are a multigene family of >25 secreted and cell surface enzymes that process or de grade various extracellular matrices65, which can be di vided into five subgroups based on substrate preference: collagenases (MMP-1, MMP-8, MMP-13), gelatinases (MMP-2, MMP-9), stromelysins (MMP-3, MMP-10), mem brane-associated MMPs (MT1-MMP, MT2-MMP), and others (e.g., MMP-12, MMP-19, MMP-20). Tissue inhibitors of metalloproteinases (TIMPs) bind to and prevent the activities of most MMPs. The relationship between pterygium and these two groups of proteins in the pathogenesis of pterygium has been studied39.
MMP-1, MMP-2, MMP-3, TIMP-1, and TIMP-3 were detected in greater amounts in pterygium tissues, epithelial cells, and fibroblasts as compared to normal conjunctiva66,67. MMP-3 was positively regulated and located in the pterygium epithelium, which may help explain the various pterygium phenotypes68. A study showed that cyclosporin A can reduce MMP-3 and MMP-13 expressions in the pterygium fibroblast culture69.
MMP and TIMP expressions vary at the different stages of pterygium. The balance break between MMPs and TIMPs may be considered to be responsible for the progression or recurrence of pterygium39.
ILs
ILs are a group of cytokines, secreted proteins, and signal molecules first seen to be expressed by the white blood cells (leukocytes). These cells play vital roles in the inflammation process; thus, ILs can be closely related to pterygium66.
The expression of IL-1α, IL-1b RA, and IL-1b precursor proteins in primary pterygium and normal conjunctival epithelium were detected via immunofluorescence. Enhanced levels of IL-1 family proteins were present in pterygium only. Likewise, IL-1α was found to be highly expressed not only in primary but also in recurrent pterygium67.
IL-6 and IL-8 were strongly expressed in the pterygium epithelium as compared to the normal cornea, conjunctiva, and limbus. In addition, IL-6 and IL-8 proteins were significantly elevated in pterygium treated with UVB, suggesting that UVB could induce the secretion of these two ILs70. IL-8 can also induce corneal vascularization directly71. IL-10 had also been reported to be expressed more in pterygium than that in the normal conjunctiva. Recently, IL-17 was found to be upregulated in the ocular surface in inflammatory pathologies, such as pterygium72.
Growth factors
A growth factor is a natural substance capable of sti mulating cellular growth, proliferation, healing, and cellular differentiation. They are important in the regula tion of various cellular processes, such as mitosis73.
Numerous growth factors are thought to have a role in pterygium pathogenesis, such as the vascular endothelial growth factor (VEGF), transforming growth factor-beta (TGF-b), basic fibroblast growth factor (bFGF), insulin-like growth factor, nervous growth factor, and connective tissue growth factor (CTGF)7. The VEGF family has been extensively investigated in ophthalmology, because of its role in pathological angiogenesis and in increasing the vascular permeability in ocular diseases, such as pterygium and retinal diseases74.
Increased expression of VEGF leads to angiogenesis and lymphangiogenesis, which may influence the normal metabolism of the connective cells and promote vascular growth. When compared to the normal conjunctiva, pterygium showed higher VEGF levels75,76. TGF-b regulates various processes common to tissue repair and disease, including fibroblast proliferation, angiogenesis, synthesis, and degradation of extracellular matrix proteins77. TGF-b1 and TGF-b2 were found to be positively regulated, whereas transforming growth factor-beta receptor 1,2 (TGFR-b1, b2) was negatively regulated in pterygium78,79.
Anti-VEGF drugs such as ranibizumab and bevacizumab have been widely used for the treatment and control of ocular diseases associated with vascular proliferation80. Although some studies suggest the use of anti-VEGF as an adjuvant therapy for surgery, studies conducted to characterize its use for the treatment of pterygium are lacking81.
Understanding the etiopathogenesis and most relevant factors involved in pterygium may allow advances on strategies to prevent its onset and progression, which may even prevent surgical procedures in the future. Although various studies have already been conducted, important genes and proteins have probably not yet been discovered. In this sense, performing additional research to better understand the etiopathogenic mechanisms and, thus, promote more targeted and effective treatment options, especially in recurrent cases, may be interesting.
REFERENCES
1. Chui J, Coroneo MT, Tat LT, Crouch R, Wakefield D, Di Girolamo N. Ophthalmic pterygium: a stem cell disorder with premalignant features. Am J Pathol. 2011;178(2):817-27.
2. Di Girolamo N, Tedla N, Kumar RK, McCluskey P, Lloyd A, Coroneo MT, et al. Culture and characterisation of epithelial cells from human pterygia [abstract]. Br J Ophthalmol. 19991;83(9):1077 LP-1082.
3. Gazzard G, Saw SM, Farook M, Koh D, Widjaja D, Chia SE, et al. Pterygium in Indonesia: prevalence, severity and risk factors. Br J Ophthalmol. 2002;86(12):1341-6.
4. Sul S, Korkmaz S, Novruzlu S. Seasonal effects on pterygium surgery outcome: implications for the role of sunlight exposure. Cornea. 2014;33(5):504-6.
5. Chui J, Girolamo N Di, Wakefield D, Coroneo MT. The pathogenesis of pterygium: current concepts and their therapeutic implications. Ocul Surf. 2008;6(1):24-43.
6. Hill JC, Maske R. Pathogenesis of pterygium. Eye (Lond). 1989;3(Pt 2): 218-26.
7. Di Girolamo N, Chui J, Coroneo MT, Wakefield D. Pathogenesis of pterygia: role of cytokines, growth factors, and matrix metalloproteinases. Prog Retin Eye Res. 2004;23(2):195-228.
8. Pyo EY, Mun GH, Yoon KC. The prevalence and risk factors for pterygium in South Korea: the Korea National Health and Nutrition Examination Survey (KNHANES) 2009-2010. Epidemiol Health. 2016;38:e2016015.
9. Song P, Chang X, Wang M, An L. Variations of pterygium prevalence by age, gender and geographic characteristics in China: A systematic review and meta-analysis. PLoS One. 2017;12(3):e0174587.
10. West S, Muñoz B. Prevalence of pterygium in Latinos: proyecto VER. Br J Ophthalmol. 2009;93(10):1287-90.
11. Anbesse DH, Kassa T, Kefyalew B, Tasew A, Atnie A, Desta B. Prevalence and associated factors of pterygium among adults living in Gondar city, Northwest Ethiopia. PLoS One. 2017;12(3):e0174450.
12. Ribeiro LA, Ribeiro LF, Castro PR, da Silva FD, Ribeiro VM, Portes AJ, et al. Characteristics and prevalence of pterygium in small communities along the Solimões and Japurá rivers of the Brazilian Amazon Rainforest. Rev Bras Oftalmol. 2011;70(6):358-62.
13. Paula JS, Thorn F, Cruz AA. Prevalence of pterygium and cataract in indigenous populations of the Brazilian Amazon rain forest. Eye (Lond). 2006;20(5):533-6.
14. Shiratori CA, Barros JC. Lourenço R. M, Padovani CR, Cordeiro R, Schellini SA [Prevalence of pterygium in Botucatu city - São Paulo State, Brazil]. Arq Bras Oftalmol. 2010;73(4):343-5. Portuguese.
15. Liu L, Wu J, Geng J, Yuan Z, Huang D. Geographical prevalence and risk factors for pterygium: a systematic review and meta-analysis. BMJ Open. 2013;3(11):e003787.
16. Rim TH, Kang MJ, Choi M, Seo KY, Kim SS. The incidence and prevalence of pterygium in South Korea: A 10-year population-based Korean cohort study. PLoS One. 2017;12(3):e0171954.
17. Viso E, Gude F, Rodríguez-Ares MT. Prevalence of pinguecula and pterygium in a general population in Spain. Eye (Lond). 2011; 25(3):350-7.
18. Shrestha S, Shrestha SM. Comparative study of prevalence of pterygium at high altitude and Kathmandu Valley. J Nepal Health Res Counc. 2014;12(28):187-90.
19. Schellini SA, dos Reis Veloso CE, Lopes W, Padovani CR, Padovani CR. Características de portadores de pterígio na região de Botucatu. Arq Bras Oftalmol. 2005;68(3):291-4.
20. Patil MS, Bawir DN, Mandlik K, Kataria I. A Study of limbal conjunctival autograft transplantation in primary pterygium. MVP J Med Sci. 2015;2(2):85.
21. Miyata K, Minami K, Otani A, Tokunaga T, Tokuda S, Amano S. Proposal for a Novel Severity Grading System for Pterygia Based on Corneal Topographic Data. Cornea. 2017;36(7):834-40.
22. Kareem AA, Farhood QK, Alhammami HA. The use of antimetabolites as adjunctive therapy in the surgical treatment of pterygium. Clin Ophthalmol. 2012;6:1849-54.
23. Sánchez-Thorin JC, Rocha G, Yelin JB. Meta-analysis on the recurrence rates after bare sclera resection with and without mitomycin C use and conjunctival autograft placement in surgery for primary pterygium. Br J Ophthalmol. 1998;82(6):661-5.
24. Fonseca EC, Rocha EM, Arruda GV. Comparison among adjuvant treatments for primary pterygium: a network meta-analysis. Br J Ophthalmol. 2018;102(6):748-56
25. Zhou WP, Zhu YF, Zhang B, Qiu WY, Yao YF. The role of ultraviolet radiation in the pathogenesis of pterygia. Mol Med Rep. 2016;14(1):3-15. Review.
26. Cooper SJ, Bowden GT. Ultraviolet B regulation of transcription factor families: roles of nuclear factor-kappa B (NF-kappaB) and activator protein-1 (AP-1) in UVB-induced skin carcinogenesis. Curr Cancer Drug Targets. 2007;7(4):325-34.
27. Kau HC, Tsai CC, Lee CF, Kao SC, Hsu WM, Liu JH, et al. Increased oxidative DNA damage, 8-hydroxydeoxy- guanosine, in human pterygium. Eye (Lond). 2006;20(7):826-31.
28. Cimpean AM, Sava MP, Raica M. DNA damage in human pterygium: one-shot multiple targets. Mol Vis. 2013;19:348-56.
29. Hou A, Lan W, Law KP, Khoo SC, Tin MQ, Lim YP, et al. Evaluation of global differential gene and protein expression in primary Pterygium: S100A8 and S100A9 as possible drivers of a signaling network. PLoS One. 2014;9(5):e97402.
30. Chalkia AK, Spandidos DA, Detorakis ET. Viral involvement in the pathogenesis and clinical features of ophthalmic pterygium. Int J Mol Med. 2013;32(3):539-43. Review.
31. Rodrigues FW, Arruda JT, Silva RE, Moura KK. TP53 gene expression, codon 72 polymorphism and human papillomavirus DNA associated with pterygium. Genet Mol Res. 2008;7(4):1251-8.
32. Tsai YY, Chang CC, Chiang CC, Yeh KT, Chen PL, Chang CH, et al. HPV infection and p53 inactivation in pterygium. Mol Vis. 2009; 15:1092-7.
33. Kelishadi M, Kelishadi M, Moradi A, Javid N, Bazouri M, Tabarraei A. human adenoviruses role in ophthalmic pterygium formation. Jundishapur J Microbiol. 2015;8(4):e16871.
34. Livezeanu C, Craitoiu MM, Manescu R, Mocanu C, Craitoiu S. Angiogenesis in the pathogenesis of pterygium. Rev Roum Morphol Embryol. 2011;52(3):837-44.
35. Young CH, Chiu YT, Shih TS, Lin WR, Chiang CC, Chou YE, et al. E-cadherin promoter hypermethylation may contribute to protein inactivation in pterygia. Mol Vis. 2010;16:1047-53.
36. Kim KW, Park SH, Kim JC. Fibroblast biology in pterygia. Exp Eye Res. 2016;142:32-9.
37. Weinstein O, Rosenthal G, Zirkin H, Monos T, Lifshitz T, Argov S. Overexpression of p53 tumor suppressor gene in pterygia. Eye (Lond). 2002;16(5):619-21.
38. Tsai YY, Chang KC, Lin CL, Lee H, Tsai FJ, Cheng YW, et al. p53 Expression in pterygium by immunohistochemical analysis: a series report of 127 cases and review of the literature. Cornea. 2005;24(5):583-6.
39. Feng QY, Hu ZX, Song XL, Pan HW. Aberrant expression of genes and proteins in pterygium and their implications in the pathogenesis. Int J Ophthalmol. 2017;10(6):973-81.
40. Ramalho FS, Maestri C, Ramalho LN, Ribeiro-Silva A, Romão E. Expression of p63 and p16 in primary and recurrent pterygia. Graefes Arch Clin Exp Ophthalmol. 2006;244(10):1310-4.
41. Tong L. Expression of p27(KIP1) and cyclin D1, and cell proliferation in human pterygium. Br J Ophthalmol. 2008;92(1):157.
42. Zhang LW, Chen BH, Xi XH, Han QQ, Tang LS. Survivin and p53 expression in primary and recurrent pterygium in Chinese patients. Int J Ophthalmol. 2011;4(4):388-92.
43. Maxia C, Perra MT, Demurtas P, Minerba L, Murtas D, Piras F, et al. Expression of survivin protein in pterygium and relationship with oxidative DNA damage. J Cell Mol Med. 2008;12(6A):2372-80.
44. Maxia C, Perra MT, Demurtas P, Minerba L, Murtas D, Piras F, et al. Relationship between the expression of cyclooxygenase-2 and survivin in primary pterygium. Mol Vis. 2009;15(15):458-63.
45. Tsujimoto Y, Finger LR, Yunis J, Nowell PC, Croce CM. Cloning of the chromosome breakpoint of neoplastic B cells with the t(14;18) chromosome translocation. Science. 1984;226(4678):1097-9.
46. Cleary ML, Smith SD, Sklar J. Cloning and structural analysis of cDNAs for bcl-2 and a hybrid bcl-2/immunoglobulin transcript resulting from the t(14;18) translocation. Cell. 1986;47(1):19-28.
47. Cui YH, Li HY, Gao ZX, Liang N, Ma SS, Meng FJ, et al. Regulation of apoptosis by miR-122 in pterygium via targeting Bcl-w. Invest Ophthalmol Vis Sci. 2016;57(8):3723-30.
48. Liu Y, Xu H, An M. mTORC1 regulates apoptosis and cell proliferation in pterygium via targeting autophagy and FGFR3. Sci Rep. 2017;7(1):7339.
49. Gumbiner BM. Cell adhesion: the molecular basis of tissue architecture and morphogenesis. Cell. 1996;84(3):345-57.
50. Beden U, Irkeç M, Orhan D, Orhan M. The roles of T-lymphocyte subpopulations (CD4 and CD8), intercellular adhesion molecule-1 (ICAM-1), HLA-DR receptor, and mast cells in etiopathogenesis of pterygium. Ocul Immunol Inflamm. 2003;11(2):115-22.
51. Kase S, Osaki M, Sato I, Takahashi S, Nakanishi K, Yoshida K, et al. Immunolocalisation of E-cadherin and beta-catenin in human pterygium. Br J Ophthalmol. 2007;91(9):1209-12.
52. Sebastiá R, Ventura MP, Solari HP, Antecka E, Orellana ME, Burnier MN. Immunohistochemical detection of Hsp90 and Ki-67 in pterygium. Diagn Pathol. 2013;8:32.
53. Liang K, Jiang Z, Ding BQ, Cheng P, Huang DK, Tao LM. Expression of cell proliferation and apoptosis biomarkers in pterygia and normal conjunctiva. Mol Vis. 2011;17:1687-93.
54. Das P, Gokani A, Bagchi K, Bhaduri G, Chaudhuri S, Law S. Limbal epithelial stem-microenvironmental alteration leads to pterygium development. Mol Cell Biochem. 2015;402(1-2):123-39.
55. Tung JN, Chiang CC, Tsai YY, Chou YY, Yeh KT, Lee H, et al. CyclinD1 protein expressed in pterygia is associated with b-catenin protein localization. Mol Vis. 2010;16:2733-8.
56. Pharmakakis N, Assimakopoulou M. Immunohistochemical detection of heat shock protein 27 and Ki-67 in human pterygium. Br J Ophthalmol. 2001;85(11):1392-3.
57. Pagoulatos D, Pharmakakis N, Lakoumentas J, Assimakopoulou M. Hypoxia-inducible factor-1α, von Hippel-Lindau protein, and heat shock protein expression in ophthalmic pterygium and normal conjunctiva. Mol Vis. 2014;20:441-57.
58. Capaldo CT, Nusrat A. Claudin switching: Physiological plasticity of the tight junction. Semin Cell Dev Biol. 2015;42:22-9.
59. Yoshida Y, Ban Y, Kinoshita S. Tight junction transmembrane protein claudin subtype expression and distribution in human corneal and conjunctival epithelium. Invest Ophthalmol Vis Sci. 2009;50(5):2103-8.
60. Dogan AS, Onder E, Arikok AT, Bicer T, Gurdal C. Claudin-1 expressions decrease in pterygium with respect to normal conjunctiva. Cutan Ocul Toxicol. 2016;35(4):315-8.
61. Pérez-Rico C, Pascual G, Sotomayor S, Montes-Mollón MA, Trejo C, Sasaki T, et al. Tropoelastin and fibulin overexpression in the subepithelial connective tissue of human pterygium. Am J Ophthalmol. 2011;151(1):44-52.
62. Zhang M, Liu Z, Xie Y. The study on the expression of keratin proteins in pterygial epithelium. Yan Ke Xue Bao. 2000;16(1):48-52.
63. Dake Y, Mukae R, Soda Y, Kaneko M, Amemiya T. Immunohistochemical localization of collagen types I, II, III, and IV in pterygium tissues. Acta Histochem. 1989;87(1):71-4.
64. Yong VW, Krekoski CA, Forsyth PA, Bell R, Edwards DR. Matrix metalloproteinases and diseases of the CNS. Trends Neurosci. 1998;21(2):75-80.
65. Sternlicht MD, Werb Z. How matrix metalloproteinases regulate cell behavior. Annu Rev Cell Dev Biol. 2001;17(1):463-516.
66. Brocker C, Thompson D, Matsumoto A, Nebert DW, Vasiliou V. Evolutionary divergence and functions of the human interleukin (IL) gene family. Hum Genomics. 2010;5(1):30-55.
67. Huang ZM, Zhang ZQ, Li YF, Du H, Yao XM. Interleukine-l alpha expression in primary and recurrent pterygium. Chin J Pract Ophthalmol. 2010;(4):407-9.
68. Seet LF, Tong L, Su R, Wong TT. Involvement of SPARC and MMP-3 in the pathogenesis of human pterygium. Invest Ophthalmol Vis Sci. 2012;53(2):587-95.
69. Kim YH, Jung JC, Jung SY, Kim YI, Lee KW, Park YJ. Cyclosporine A downregulates MMP-3 and MMP-13 expression in cultured pterygium fibroblasts. Cornea. 2015;34(9):1137-43.
70. Di Girolamo N, Kumar RK, Coroneo MT, Wakefield D. UVB-mediated induction of interleukin-6 and -8 in pterygia and cultured human pterygium epithelial cells. Invest Ophthalmol Vis Sci. 2002; 43(11):3430-7.
71. Koch AE, Polverini PJ, Kunkel SL, Harlow LA, DiPietro LA, Elner VM, et al. Interleukin-8 as a macrophage-derived mediator of angiogenesis. Science. 1992;258(5089):1798-801.
72. Jabarin B, Solomon A, Amer R. Interleukin-17 and its correlation with vascular endothelial growth factor expression in ocular surface pathologies: a histologic study. Eur J Ophthalmol. 2016; 26(4):283-6.
73. Tyson JJ, Novak B. Control of cell growth, division and death: information processing in living cells. Interface Focus. 2014;4(3):20130070.
74. Witmer AN, Vrensen GF, Van Noorden CJ, Schlingemann RO. Vascular endothelial growth factors and angiogenesis in eye disease. Prog Retin Eye Res. 2003;22(1):1-29.
75. Fukuhara J, Kase S, Ohashi T, Ando R, Dong Z, Noda K, et al. Expression of vascular endothelial growth factor C in human pterygium. Histochem Cell Biol. 2013;139(2):381-9.
76. Gumus K, Karakucuk S, Mirza GE, Akgun H, Arda H, Oner AO. Overexpression of vascular endothelial growth factor receptor 2 in pterygia may have a predictive value for a higher postoperative recurrence rate. Br J Ophthalmol. 2014;98(6):796-800.
77. Govinden R, Bhoola KD. Genealogy, expression, and cellular function of transforming growth factor-beta. Pharmacol Ther. 2003; 98(2):257-65.
78. Zhong M, Shen W, Fu Q, Zhang Y. Quantitative detection of the expression level of transforming growth factor beta and its receptors in pterygium with RT- PCR. Int J Ophthalmol. 2009;2(4):302-5.
79. Bianchi E, Scarinci F, Grande C, Plateroti R, Plateroti P, Plateroti AM, et al. Immunohistochemical profile of VEGF, TGF-b and PGE2 in human pterygium and normal conjunctiva: experimental study and review of the literature. Int J Immunopathol Pharmacol. 2012; 25(3):607-15.
80. Hoyama E, Viveiros MM, Shiratori C, de Oliveira DE, Padovani CR, Selva D, et al. Expression of vascular endothelial growth factor (VEGF) in macrophages, fibroblasts, and endothelial cells in pterygium treated with 5-Fluorouracil. Semin Ophthalmol. 2015; 30(3):171-6.
81. Zeng W, Liu Z, Dai H, Yan M, Luo H, Ke M, et al. Anti-fibrotic, anti-VEGF or radiotherapy treatments as adjuvants for pterygium excision: a systematic review and network meta-analysis. BMC Ophthalmol. 2017;17(1):211.
Submitted for publication:
November 12, 2018.
Accepted for publication:
May 28, 2019.
Funding: This study received no specific financial support.
Disclosure of potential conflicts of interest: None of the authors have any potential conflicts of interest to disclose.
How to cite this article:
 ABO is licensed under a Creative Commons Attribution-NonComercial 4.0 Internacional.
ABO is licensed under a Creative Commons Attribution-NonComercial 4.0 Internacional.